Oxygen is evolved from the splitting of water in the photosystem II (PSII) complex, where light energy is first absorbed and electrons resulting from this split are used for photochemistry. PS II activity can be monitored with a chlorophyll fluorometer. Pea plants (Pisum sativum) were studied in controlled environments and in the field, under varying light and temperature. PS II yield (PhiPSII) and oxygen evolution (JO2) were measured. Light response curves were generated and maximum quantum yield and maximum rate of oxygen evolution were determined. Temperature (20-35oC) had no effect on the rate of O2 evolution at low light, while increasing temperature stimulated O2 evolution under high light levels. Maximum quantum yields for O2 evolution under limiting light were 70-80% of the theoretical maximum value based on the quantum requirements through the two photosystems. Near maximum rates of O2 evolution occurred at about 40% of full sunlight (800 (mol quanta m-2 s-1). Above this light level, there is inefficient use of light which could limit crop productivity.
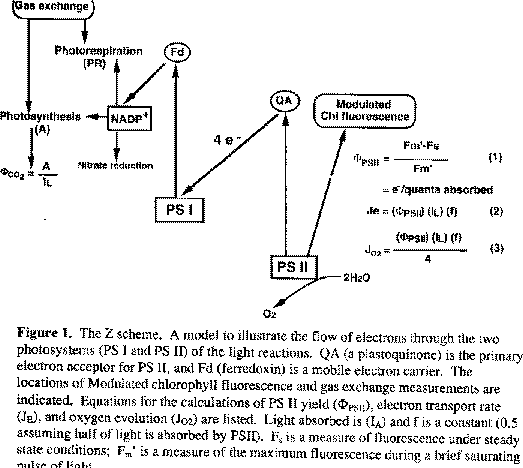
Photosynthesis is the foundation for life on this planet. Through the reactions of photosynthesis the sunís energy is harvested, converting inorganic molecules to organic molecules that can be utilized by all other forms of life. Photosynthesis consists of light and dark reactions. In the light reactions, photons of light are absorbed, and a strong reductant is generated using electrons which have been split from water. This reductive power is then used to drive electron transfer reactions which eventually produce ATP and NADPH, that serve as assimilatory power for the dark reactions. The flow of electrons through photosystems I and II of the light reactions is often portrayed in the Z scheme pathway (Figure 1). CO2 is reduced in the dark reactions, reacting with ribulose 1,5-bisphosphate (RuBP) catalyzed by ribulose 1,5-bisphosphate carboxylase (Rubisco) in the first reaction of the C3 cycle. Through this cycle, CO2 is transformed into carbohydrates.
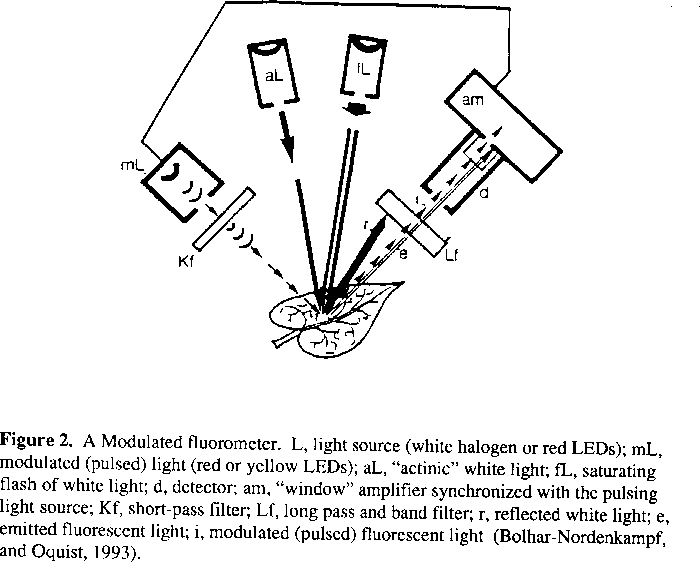
Traditionally photosynthesis has been measured in a gas exchange cuvette, however this method confines the leaf to an artificial environment. Recent evidence suggests that photosynthesis can also be measured via a novel technique using a fluorometer to analyze chlorophyll a fluorescence emitted by the leaf after light absorption. Fluorescence analysis gives a measure of the activity of photosystem II (PSII). Measuring photosynthesis in this manner is beneficial because it provides a non-invasive method to monitor the harvesting of light energy by the chloroplast in vivo (Figure 2). Chlorophyll fluorescence can also be used as a quick screen for the response of plants which have been under stress such as water or salinity stresses (Krause, 1991).
Oxygen is evolved from the splitting of water in the initial steps of photosynthesis. Water is cleaved into its component parts: H+, e- and O2 and the electrons are then sent through the light reaction pathway. For each (mol of O2 evolved, a minimum of eight (mol of quanta must be absorbed, four by each photosystem to drive the transfer of four electrons through the pathway (Figure 1). This results in a theoretical maximum quantum yield for O2 evolution (PhiO2) of 0.125 mol of O2 per mol of quanta. The efficiency of PS II in producing oxygen can be determined and compared to the above theoretical considerations. I investigated oxygen evolution and photosystem II yield (PhiPSII) in Pisum sativum leaves using a modulated fluorometer in a pea field canopy and in the laboratory on plants grown in a greenhouse.
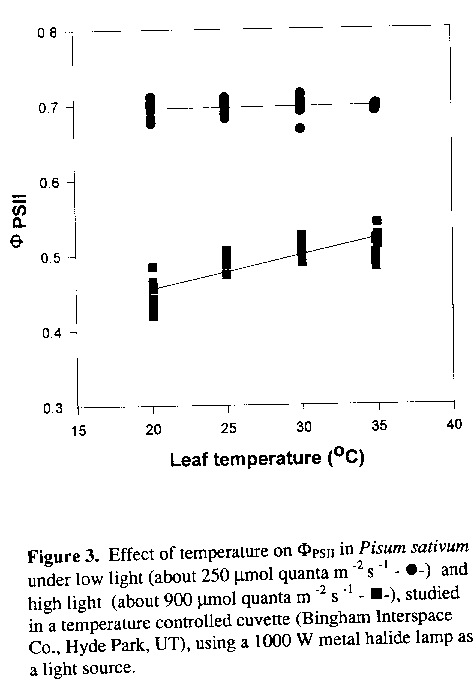
Since both light and temperature are factors affecting photosynthesis, the two were observed separately to determine how much each affected photosynthetic yield (PhiPSII) within the parameters of this study. Changing temperature between 20 to 35oC had little or no affect on PhiPSII. At low light, yield is independent of temperature and at high light there was only a slight increase in yield as temperature increased (Figure 3). This suggests that at low light the rate of oxygen evolution, which is calculated using PhiPSII as a factor, is not affected by temperature. Only at higher light levels does temperature begin to influence this rate, but the effect is small compared to the effect of changing light levels. Since temperature, between 20 and 35oC, had little affect on PhiPSII, these results suggest that temperature over this range does not notably affect oxygen evolution in Pisum sativum.
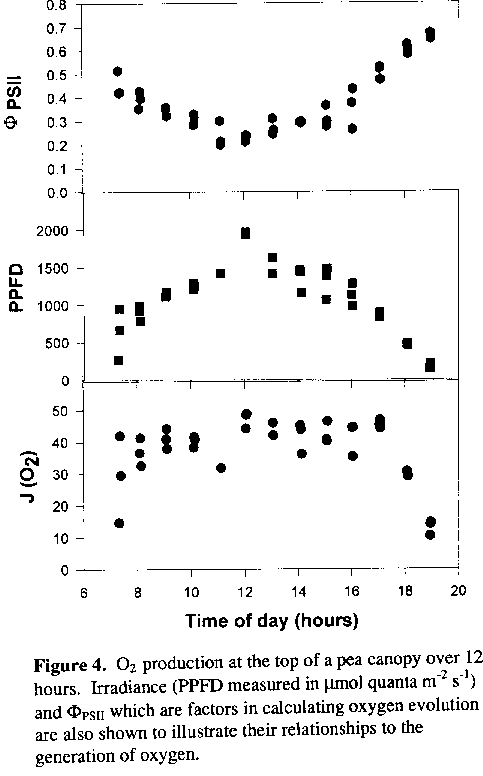
Light response curves for PhiPSII and JO2 were generated from measurements on laboratory and field plants. Field measurements included analysis from three levels in the canopy (top, middle, and bottom) on five individual plants during the course of the day. Some representative data are presented (Figure 4). The maximum yield for photosynthetic O2 evolution (PhiO2), the maximum rate of oxygen evolution (JO2 max ) and the average rate of oxygen evolved (JO2 ) were calculated (Table 1).
Theoretically, a minimum of one (mol of O2 is evolved for every eight (mol of quanta absorbed, assuming a minimum of four quanta per photosystem. This means that the theoretical maximum rate of oxygen evolution per quanta is 1/8 or 0.125. The yield may be lower if some of the absorbed quanta are lost as heat or fluorescence. The maximum experimental quantum yield to O2 evolution is best measured under low light when the highest percentage of absorbed quanta are used for photochemistry. The results indicate that the experimental maximum yields are about 70-80% of the theoretical maximum. This is reasonable considering that some energy loss occurs under limiting light due to fluorescence and heat; thus light harvesting is not 100% efficient. These results also compare favorably with the maximum PhiO2 values of 0.103 obtained by Bjorkman and Demmig (1987) using an O2 electrode measuring system. The slightly lower values of PhiO2 in the middle and top leaves of the canopy could be due to photoinhibition occurring during periods of excess light in the upper canopy (Table 1 and Figure 5).
It was also found that leaves receiving light levels of more than 800 PPFD can not make efficient use of this additional light. This implies that a factor other than light is limiting the rate of oxygen evolution at higher intensities. The rate of oxygen evolution has been shown to be related to the electron transport capacity and elements of the electron transport chains (Evans, 1987). PS II activity and CO2 fixation have also been shown to be linked (Oberhuber, et al.) . These factors may indicate that an element of the electron transport chain or Rubisco, ribulose 1,5-bisphosphate or another factor in the Calvin cycle is the limiting factor for oxygen evolution in peas at light intensities greater than 800 PPFD.
The maximum JO2 values under full sunlight are lower for the bottom leaves of the canopy, suggesting that these leaves more limiting in components of electron transport or carbon fixation. As shown in Table I, JO2 values over the course of the day increase from the bottom to the top of the canopy. This occurs because the upper leaves receive more light than the lower leaves. However, the upper leaves are not functioning at maximum efficiency under high light which could limit productivity. Figure 4 illustrates that an upper leaf in the canopy maintains a constant rate of photosynthesis (JO2) over much of the photoperiod. However, it also shows that the quantum yield of PSII (PhiPSII) drops as light intensity increases to a maximum value at midday, indicating that the upper leaves are not functioning at maximum efficiency under high light This inefficiency is also reflected in light saturation of photosynthesis at about 40% of full sunlight which could limit crop productivity.
I gratefully acknowledge Gerald Edwards, Raymon Donahue, and Dongxiang He for their advice and guidance. This project was supported by the Plant Biochemistry Training Grant funded by the Department of Energy.
Bjorkman, O. and B. Demmig. (1987). Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta. 170: 489-504.
Bolhar-Nordenkampf, H. R. and G. Oquist. 1993. Chlorophyll fluorescence as a tool in photosynthesis research. In: Photosynthesis and Production in a Changing Environment: a field and laboratory manual. (Hall, D. O., Scurlock, J. M. O., Bolhar-Nordenkampf, H. R., Leegood, R. C. and Long, S. P., eds). pp. 193-206. Chapman and Hall, London.
Evans, J. R. 1987. The relationship between electron transport components and photosynthetic capacity in pea leaves grown at different irradiances. Aust. J. Plant Physiol. 14: 157-70.
Evans, J. R., I. Jakobsen, and E. Ogren. 1993. Photosynthetic light-response curves: Gradients of light absorption and photosynthetic capacity. Planta. 189: 191- 200.
Genty, B, J. M. Briantais, and N. R. Baker. 1989. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim Biophys Acta. 990: 87-92.
Krause, G. H. and E. Weis. 1991. Chlorophyll fluorescence and photosynthesis: The basics. Annu. Rev. Plant Physiol. Plant Mol. Biol. 42: 313-49.
Oberhuber, W. and G. E. Edwards. 1992. Quantum yield of photosystem II and CO2 fixation in C3 and C4 plants. Research in Photosynthesis. 3: 907-910.
Oberhuber, W., Z. Dai, G. E. Edwards. 1993. Light dependence of quantum yields of Photosystem II and CO2 fixation in C3 and C4 plants. Photosynthesis Research. 35: 265-274.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}